
Cannibalism in pre-history: taboo or standard practice?
- Shruti Kamath

- Oct 23, 2022
- 11 min read
Anthropophagy or human cannibalism continues to spark a morbid curiosity in modern societies. The topic has traditionally given rise to fascination or revulsion – either of which is usually induced by cultural bias or ignorance. Today, cannibalism is often likened to barbarism, and is a taboo in most societies, evoking ethical and moral arguments. However, a lot of ethnographic and anthropological data demonstrates that this behaviour was not unusual in human history and prehistory, and has been used for a variety of reasons over time. Indeed, the motivations and social contexts in which this behaviour occurs are often linked to complex and abstract ideals. Here, we consider the case for prehistoric cannibalism, and why it may have been adopted by early humans.

Anthropophagy (from the Greek word: anthrōpos "man; human being", and stem phagein: “to eat”) is the act or custom of eating human flesh. The term “cannibalism’’ has a more colonial etymology - the Spanish accused the people of the Caribbean of ritualistically eating their enemies, and thus called these indigenous people - “Caníbales”. Cannibalism is a universal act, seen in different species, which use it as a means of population control, a source of food, or a way of showing authority and strength. While the term “cannibalism” is usually considered derogatory, it may be used by an individual or group for several reasons.
Ethnographic studies have suggested the presence of different types of cannibalism, including:
Survival cannibalism: A Uruguayan rugby team whose plane crashed in the Andes in 1972 while travelling for a match to Chile, resorted to eating their dead teammates in order to survive.
Nutritional or gastronomic: In ancient China, it was not unusual for emperors and members of the royal families to enjoy human body parts as part of their meal
Ritual or funerary: Pano Kaxinawá, an indigenous tribe from Brazil, practises this form of cannibalism, where, consuming the body of a loved one is considered an act of compassion and self-protection, and would ensure the rapid release of the individual’s soul. (d) Warfare: The Wari tribes from Peru cannibalise members of other groups, where enemies’ bodies are treated with less respect than those of animals, and large parts of bodies are wasted
Medicinal: From the 17th to 19th century, human fat was sold in German markets as a remedy for arthritis, broken bones, and sprains
Psychotic or criminal: an example would be the American serial killer Jeffrey Dahmer who ate his victims. Apart from the different purposes, we can further classify cannibalism depending on the origin of the people being consumed:
Endo-cannibalism: where the consumed is part of the same family or social group
Exo-cannibalism: where the consumed is part of a different group.

Hence, cannibalism can be practised for a variety of reasons, and has been by hominins (the group comprising modern humans, and all our immediate ancestors) from the Early Pleistocene (EP) (2.5 mya [million years ago] to 781 kya [thousand years ago]), to the present times. While cannibalism is a controversial topic today, it is even more so for anthropologists dealing with prehistoric fossils. There is an undeniable link with cannibalism and the cultural systems of a group – the social beliefs, traditions, and their way of life. Here lies the problem with the prehistoric record - while it is possible to identify signs of cannibalism in an archaeological site, it may not always be possible to extrapolate the ‘why’, as some of these expressions would be archaeologically unrecognisable. Nonetheless, there is a record for such practices among different hominin species, including Homo antecessor (EP, age range of 779 – 949 kya), Homo neanderthalensis or Neanderthals (around 400 – 40 kya), as well as in Homo sapiens or modern humans - from as early as the Upper Palaeolithic (50 kya – 11 kya) onwards, to the present times.
Cannibalism in prehistory is identified through certain taphonomic (study dealing with the processes of fossilisation) criteria, and this is further used to deduce the probable cause for this behaviour. First, the signs that indicate that an individual’s remains have been intentionally exploited by hominins for consumption, include: fractured bones, percussion pits (the impact point where a stone or any solid material was intentionally struck on the bone cortex, scarring the surface), tool-induced surface modifications such as cut marks (for removal of ligament and muscle attachments, defleshing, skinning), chop marks (cutting larger muscle attachments/dismembering), scraping marks (for muscle removal), and tooth marks.
When trying to identify the purpose, for example, nutritional versus ritualistic cannibalism, certain criteria may be adopted, although these may not always be clear-cut. For example, for nutritional cannibalism, the hominin and animal remains are butchered in the same fashion, the frequency and type of cut marks are similar, and the hominin and animal remains may even be found together. However, in most cases with ritualistic or funerary cannibalism, the cut marks may vary in size and intensity than those of the animal bones, and usually, the hominin remains are found separate from the animal bones.
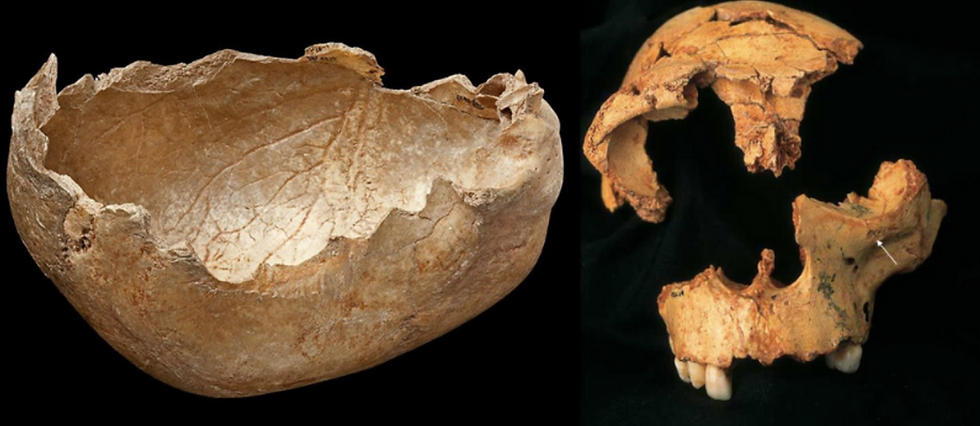
The earliest known case of cannibalism comes from the EP archaeological site of Gran Dolina (TD6), Sierra de Atapuerca, Spain. This site has been dated to 779 – 949 kya, with over 160 hominin fossil remains, and a minimum number of nine individuals, and have been identified as the species Homo antecessor. The age of death of these individuals is of particular interest, as they range from immature individuals to young adults. About 44.5% of these remains show some sign of hominin-induced bone modification, including cut marks, bone breakage, and even tooth marks. The remains are mixed with a large number of faunal remains, suggesting that this was not a time of food stress, and hence, a case of nutritional cannibalism. Another study approached this problem through the comparative analysis of chimpanzee behaviour. For example, chimpanzees (who are useful models for understanding hominin behaviour) can partake in inter-group attacks, especially during times of abundant food resources, concluding in lethal results, such as exocannibalism of infants. This would increase their catchment territory, along with a larger pool of fertile females, and greater resources. Thus, it is highly plausible that Homo antecessor practised exo-cannibalism, perhaps for the purpose of increasing their catchment territory. The consumed were either immature or young adults – individuals who would have posed the least risk in their capture. The archaeological data indicate that the cannibalism at TD6 occurred frequently over dozens, if not, over hundreds of years, forming part of their culture/subsistence patterns, and that the hominin remains did not receive any special treatment or symbolic burial. Hence, the cannibalism could have been nutritional.
In the Middle Pleistocene (MP; 781 – 126 kya) site of Arago, Tautavel, France, over 148 hominin remains have been found, representing 18 adults and 12 juveniles. It has been proposed that these hominins belong to Homo heidelbergensis, however, the debate continues regarding the validity of this species. Most of the fossil remains show some levels of systematic breakage, with an abundance of cut marks on the long bones and skulls. It is interesting to note that only skulls, mandibles and long limb bones have been found. The lack of trunk, hand and foot bones, along with a variable distribution of the hominin bones from those of the other faunal remains, may indicate a ritualistic purpose. However, considering the general poor survival rate of these bones, and the issue of identifying smaller bones in a fragmented assemblage, it is likely that this very well may have been a case of nutritional, rather than ritualistic cannibalism.
Across the MP and Late Pleistocene (LP; 125 – 11 kya), we see the appearance, and the gradual demise of the Neanderthals, and along with it, multiple evidence for cannibalism in the archaeological record. The Krapina Cave site, in Croatia, has around 900 Neanderthal remains, and dates back to 120 – 130 kya. The case for cannibalism at Krapina has been disputed by some; however, the bones demonstrate cut marks that are different from the remains of reindeer found at the site, which may imply a ritualistic behaviour, such as - preparing and cleaning bones for burial. Others believe the remains show aggressive cut marks, and even tooth marks, suggestive of nutritional cannibalism. The El Sidrón cave, Asturias, Spain, dating to around 49 kya, has over 2000 Neanderthal remains, 10% of which show some form of hominin-induced damage. These bones are found alongside stone tools, with minimal representation of faunal remains. The lack of fauna corresponds to periods of nutritional stress, and may indicate compensation for food shortage periods. The methods by which the remains were processed (harsh modification methods) further supports this. Ultimately, the low (less than ten sites) geographical and chronological evidence for Neanderthal cannibalism, along with the methods used (higher frequency of cut marks compared to animal bones), could potentially suggest that this was not common practice in this species.
In the Upper Palaeolithic site of Gough’s Cave (12,600 BCE), Somerset, England, human and non-human remains were found discarded in the same pit, suggestive of gastronomic cannibalism; the skulls were modelled and fashioned as skull cups. In the Mesolithic site of Brillenhöhle (13,170 - 12,450 BCE), Germany, a set of human remains, with cut marks, and tooth marks, suggest gastronomic cannibalism was practised here. In the Neolithic site of Fontbrégoua (3,500 - 2,850 BCE), France, 13 human remains show clear signs of butchery, similar to that of faunal remains; some remains lack skulls, and it was assumed that this was a form of warfare cannibalism; but it has later been hypothesised that this was a form of funerary ritual.
When trying to identify the purpose, for example, nutritional versus ritualistic cannibalism, certain criteria may be adopted, although these may not always be clear-cut. For example, for nutritional cannibalism, the hominin and animal remains are butchered in the same fashion, the frequency and type of cut marks are similar, and the hominin and animal remains may even be found together. However, in most cases with ritualistic or funerary cannibalism, the cut marks may vary in size and intensity than those of the animal bones, and usually, the hominin remains are found separate from the animal bones.
The earliest known case of cannibalism comes from the EP archaeological site of Gran Dolina (TD6), Sierra de Atapuerca, Spain. This site has been dated to 779 – 949 kya, with over 160 hominin fossil remains, and a minimum number of nine individuals, and have been identified as the species Homo antecessor. The age of death of these individuals is of particular interest, as they range from immature individuals to young adults. About 44.5% of these remains show some sign of hominin-induced bone modification, including cut marks, bone breakage, and even tooth marks. The remains are mixed with a large number of faunal remains, suggesting that this was not a time of food stress, and hence, a case of nutritional cannibalism. Another study approached this problem through the comparative analysis of chimpanzee behaviour. For example, chimpanzees (who are useful models for understanding hominin behaviour) can partake in inter-group attacks, especially during times of abundant food resources, concluding in lethal results, such as exocannibalism of infants. This would increase their catchment territory, along with a larger pool of fertile females, and greater resources. Thus, it is highly plausible that Homo antecessor practised exo-cannibalism, perhaps for the purpose of increasing their catchment territory. The consumed were either immature or young adults – individuals who would have posed the least risk in their capture. The archaeological data indicate that the cannibalism at TD6 occurred frequently over dozens, if not, over hundreds of years, forming part of their culture/subsistence patterns, and that the hominin remains did not receive any special treatment or symbolic burial. Hence, the cannibalism could have been nutritional.
In the Middle Pleistocene (MP; 781 – 126 kya) site of Arago, Tautavel, France, over 148 hominin remains have been found, representing 18 adults and 12 juveniles. It has been proposed that these hominins belong to Homo heidelbergensis, however, the debate continues regarding the validity of this species. Most of the fossil remains show some levels of systematic breakage, with an abundance of cut marks on the long bones and skulls. It is interesting to note that only skulls, mandibles and long limb bones have been found. The lack of trunk, hand and foot bones, along with a variable distribution of the hominin bones from those of the other faunal remains, may indicate a ritualistic purpose. However, considering the general poor survival rate of these bones, and the issue of identifying smaller bones in a fragmented assemblage, it is likely that this very well may have been a case of nutritional, rather than ritualistic cannibalism.
Across the MP and Late Pleistocene (LP; 125 – 11 kya), we see the appearance, and the gradual demise of the Neanderthals, and along with it, multiple evidence for cannibalism in the archaeological record. The Krapina Cave site, in Croatia, has around 900 Neanderthal remains, and dates back to 120 – 130 kya. The case for cannibalism at Krapina has been disputed by some; however, the bones demonstrate cut marks that are different from the remains of reindeer found at the site, which may imply a ritualistic behaviour, such as - preparing and cleaning bones for burial. Others believe the remains show aggressive cut marks, and even tooth marks, suggestive of nutritional cannibalism. The El Sidrón cave, Asturias, Spain, dating to around 49 kya, has over 2000 Neanderthal remains, 10% of which show some form of hominin-induced damage. These bones are found alongside stone tools, with minimal representation of faunal remains. The lack of fauna corresponds to periods of nutritional stress, and may indicate compensation for food shortage periods. The methods by which the remains were processed (harsh modification methods) further supports this. Ultimately, the low (less than ten sites) geographical and chronological evidence for Neanderthal cannibalism, along with the methods used (higher frequency of cut marks compared to animal bones), could potentially suggest that this was not common practice in this species.
In the Upper Palaeolithic site of Gough’s Cave (12,600 BCE), Somerset, England, human and non-human remains were found discarded in the same pit, suggestive of gastronomic cannibalism; the skulls were modelled and fashioned as skull cups. In the Mesolithic site of Brillenhöhle (13,170 - 12,450 BCE), Germany, a set of human remains, with cut marks, and tooth marks, suggest gastronomic cannibalism was practised here. In the Neolithic site of Fontbrégoua (3,500 - 2,850 BCE), France, 13 human remains show clear signs of butchery, similar to that of faunal remains; some remains lack skulls, and it was assumed that this was a form of warfare cannibalism; but it has later been hypothesised that this was a form of funerary ritual.

In the Bronze Age site of El Mirador (2,450 - 2150 BCE), Atapuerca, Spain, six skull cups have been found; this, along with other remains showing cut marks, bone breakage, and tooth marks, indicate gastronomic cannibalism.
The study of prehistoric cannibalism has two objectives - to determine whether the skeletal remains truly show signs of cannibalism, and to determine the causes for these verified cases. However, in most cases, it is possible to identify the signs and contexts, but not causes for it. The historic and ethnographic cannibalism that are known today took place in structured, hierarchical societies, with high symbolisms and myths associated with them. Cannibalism is but a part of this structured world. The Pleistocene hominins may have adopted this behaviour due to a variety of reasons – from environmental stresses, to cultural practices, or a combination of factors. There is no way of truly comprehending these behaviours, and hence, our clear-cut classifications for why cannibalism occurs, may not apply to these people.
Further reading:
Edwards, P., 2015. 7 surprising facts about cannibalism. Vox. https://www.vox.com/2015/2/17/8052239/cannibalism-surprising-facts.
Saladié, P. and Rodríguez-Hidalgo, A., 2017. Archaeological evidence for cannibalism in prehistoric Western Europe: From Homo antecessor to the bronze age. Journal of Archaeological Method and Theory, 24(4), pp.1034-1071.
Saladie, P., Huguet, R., Rodriguez-Hidalgo, A., Caceres, I., Esteban-Nadal, M., Arsuaga, J.L., de Castro, J.M.B. and Carbonell, E., 2012. Intergroup cannibalism in the European Early Pleistocene: The range expansion and imbalance of power hypotheses. Journal of Human Evolution, 63(5), pp.682-695.
Yustos, M. and de los Terreros, J.Y.S., 2015. Cannibalism in the Neanderthal world: an exhaustive revision. Journal of taphonomy, 13(1), pp.33-52.
Worrall, S., 2016. After the plane crash—and the cannibalism—a life of hope. National Geographic. Retrieved from https://www. nationalgeographic. com/news/2016/04/160403-andes-uruguay-rugby-cannibal-plane-crash-canessa-ngbooktalk.
Photo credit:
Photo credit: Nikola Solic/Reuters/Corbis. Sourced from https://www.theguardian.com/science/2017/apr/06/prehistoric-cannibalism-not-just-driven-by-hunger-study-reveals Dorey,
Javier Trueba/Madrid Scientific Films. Source from https://australian.museum/learn/science/human-evolution/homo-antecessor/.
Natural History Museum

Comments